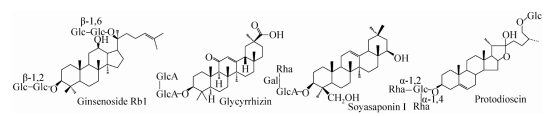
Les saponines sont une classe de glycosides dont les aglycones sont des composés triterpéniques ou stériques. Elles constituent l'un des ingrédients efficaces de nombreuses plantes médicinales chinoises telles que le ginseng, la réglisse et l'igname (la structure principale des saponines est présentée à la figure 1). Renforcer l'immunité et d'autres fonctions. Il existe de nombreux rapports sur la biotransformation des ginsénosides dans la littérature. À l'heure actuelle, plus de 150 types de ginsénosides ont été séparés et identifiés. Les teneurs en ginsénosides Rb1, Rb2, Rc, Rd, Re et Rg1 atteignent 80%, tandis que les teneurs en ginsénosides Rg3, Rh2, F2, composé K (C-K) et autres saponines rares sont faibles ou inexistantes. Des études ont montré que certaines saponines rares ont de bonnes activités pharmacologiques. Toutefois, en raison de leur faible teneur, leur préparation et leur production sont limitées. Le même type de ginsénoside possède le même aglycone, mais la chaîne de sucre est différente. Les ginsénosides rares et la teneur plus élevée du même type de saponines ne diffèrent souvent que par 2 ou 3 groupes de sucres. Par conséquent, le même type de saponine rare active peut être préparé par hydrolyse enzymatique de la saponine à haute teneur.
Figure 1. Structure des principales saponines
Les différentes glycosides hydrolases ont des sélectivités différentes et les voies d'hydrolyse des ginsénosides sont également différentes. Comme le montre le tableau 1, différentes glycosides hydrolases peuvent être utilisées pour préparer différents ginsénosides rares. Le ginsénoside Rd peut être préparé en hydrolysant les ginsénosides Rb1, Rb2, Rb3 et le groupe de sucre externe C-20 de Rc. La β-glucosidase isolée et purifiée de l'escargot de jade blanc de Chine et de Thermus caldophilus peut convertir le ginsénoside Rb1 en Rd. Kim et al. l'ont obtenue à partir de micro-organismes du sol en utilisant la technologie du clonage moléculaire pour convertir le ginsénoside Rb1, qui est la glycoside hydrolase recombinante du Rd. Par la suite, les chercheurs ont cloné la glucosidase de Thermotoga thermarum et de Bifidobacterium longum H-1, ce qui a amélioré l'efficacité de la transformation et de la préparation du ginsénoside Rd. La glucosidase obtenue à partir de Flavobacterium johnsoniae et Thermus thermophilus par technologie recombinante peut non seulement convertir le ginsénoside Rb1 en Rd, mais aussi hydrolyser la chaîne de sucre C-20 du gypénoside XVII (G17) pour produire le ginsénoside F2. Outre la glucosidase, l'α-L-arabinofuranoside hydrolase qui peut convertir le ginsénoside Rc en Rd a été obtenue à partir de racines de ginseng et de Leuconostoc sp. La Α-L-arabinofuranoside hydrolase et l'α-L-arabinopyranoside hydrolase sont obtenues à partir de Bifidobacterium breve et Bifidobacterium longum, qui peuvent transformer le ginsénoside Rc et Rb2 en Rd. Il a été rapporté dans la littérature que l'α-L-arabinofuranoside hydrolase de Caldicellulosiruptor saccharolyticus et Rhodanobacter ginsenosidimutans peut non seulement hydrolyser le ginsénoside Rc en Rd, mais aussi convertir le composé Mc1 (C-Mc1) en F2. La glycoside hydrolase isolée et purifiée à partir d'Aspergillus par Yu et al. peut convertir tous les ginsénosides Rb1, Rb2, Rb3 et Rc en Rd. Certaines glycosides hydrolases peuvent hydrolyser complètement les chaînes de sucre en position C-20 dans des molécules telles que les ginsénosides de type glycol Rb1, Rb2, Rb3, Rc et Rd, pour générer le ginsénoside Rg3, qui permet une production à grande échelle de Rg3 et est développé en tant que médicament anti-tumoral. La glucosidase de Paecilomyces bainier et de Microbacterium esteraromaticum peut hydrolyser directement le ginsénoside Rb1 en Rg3, tandis que la glucosidase isolée et purifiée de Microbacterium esteraromaticum peut hydrolyser le ginsénoside Rb2 en Rg3. La glycoside hydrolase recombinante clonée à partir de Pseudonocardia par la technologie du clonage moléculaire peut transformer les ginsénosides Rb1, Rb3 et Rd pour préparer le Rg3. De même, une série de ginsénosides rares et actifs peut être préparée en hydrolysant le groupe de sucre en position C-3 dans les ginsénosides. La glucosidase recombinante clonée à partir de Sphingomonas et de Sphingopyxis alaskensis peut hydrolyser le glucose à l'extérieur de la chaîne de sucre en position C-3 dans les molécules de ginsénoside Rb1, Rb2, Rc, Rd et Rg3, et préparer le G17, le composé O (CO) et les C-Mc1, F2 et Rh2. Certaines glycosidases peuvent hydrolyser directement le groupe glucosyle interne en position C-3. Par exemple, la glucosidase de Terrabacter ginsenosidimutans et Esteya vermicola peut hydrolyser la chaîne de sucre en position C-3 des molécules de ginsénoside Rb1, Rb2, Rb3, Rc et Rd pour produire la saponine correspondante LXXV (G75), le composé Y (C-Y), le composé Mx (C-Mx), le composé Mc (C-Mc) et C-K. En outre, certaines glycosides hydrolases peuvent hydrolyser simultanément les groupes de sucre C-20 et C-3 dans les ginsénosides de type glycol. La glucosidase recombinante clonée à partir de l'Arthrobacter chlorophenolicus peut convertir les ginsénosides Rb1, Rb2 et Rc en F2. La glycoside hydrolase de Fusobacterium K60, des champignons endophytes GE 17-18, de Sulfolobus acidocaldarius, d'Aspergillus niger et de Microbacteriu esteraromaticum peut hydrolyser le ginsénoside Rb1 pour produire C-K.
Tableau 1. Biotransformation des ginsénosides par les glycosidases
Produit
Substrat
Réaction
Organisme
Rd
Rb1
β-Glucosidase
Escargot de jade blanc de Chine
Rd
Rb1
β-Glucosidase
Thermuscaldophile
Rd
Rb1
β-Glucosidase
Bactéries non cultivées
Rd
Rb1
β-Glucosidase
Thermotogathermarum
Rd
Rb1
β-Glucosidase
BifidobacteriumlongumH-1
Rd
Rb1
β-Glucosidase
Flavobacteriumjohnsoniae
Rd
Rb1
β-Glucosidase
Thermusthermophilus
Rd
Rb1
β-Glucosidase
Penicilliumoxalicum
Rd
Rb1
β-Glucosidase
Cladosporiumfulvum
Rd
Rc
α-L-Arabinofuranosidase
Panaxginseng
Rd
Rc
α-L-Arabinofuranosidase
Leuconostoc
Rd
Rc
α-L-Arabinofuranosidase
Bifidobacteriumbref
Rd
Rc
α-L-Arabinofuranosidase
Bifidobacteriumlongum
Rd
Rc
α-L-Arabinofuranosidase
Caldicellulosiruptorsaccharolyticus
Rd
Rc
α-L-Arabinofuranosidase
Rhodanobacterginsenosidimutans
Rd
Rb2
α-L-Arabinopyranosidase
Bifidobacteriumbref
Rd
Rb2
α-L-Arabinopyranosidase
Bifidobacteriumlongum
Rd
Rb1/Rb2/Rb3/Rc
Glycosidase
Aspergillus
Rg3
Rb1
β-Glucosidase
Paecilomycesbainier
Rg3
Rb1
β-Glucosidase
Microbactériesesteraromaticum
Rg3
Rb2
β-Glucosidase
Microbactériesesteraromaticum
Rg3
Rb1/Rb3/Rd
β-Glucosidase
Pseudonocardia
G17
Rb1
β-Glucosidase
Sphingomonas
G17
Rb1
β-Glucosidase
Sphingopyxisalaskensis
G17
Rb1
β-Glucosidase
Cellulosimicrobiumcelluloses
G75
Rb1
β-Glucosidase
Terrabacterginsenosidimutans
G75
Rb1
β-Glucosidase
Esteyavermicola
F2
G17
β-Glucosidase
Flavobacteriumjohnsoniae
F2
G17
β-Glucosidase
Thermusthermophilus
F2
C-Mc1
α-L-Arabinofuranosidase
Caldicellulosiruptorsaccharolyticus
F2
C-Mc1
α-L-Arabinofuranosidase
Rhodanobacterginsenosidimutans
F2
Rd
β-Glucosidase
Cellulosimicrobiumcelluloses
F2
Rb1/Rb2/Rc
β-Glucosidase
Arthrobacterchlorophénolique
Rh2
Rg3
β-Glucosidase
Sphingopyxisalaskensis
CK
Rd
β-Glucosidase
Terrabacterginsenosidimutans
CK
Rd
β-Glucosidase
Esteyavermicola
CK
Rb1
β-Glucosidase
FusobacteriumK-60
CK
Rb1
β-Glucosidase
champignons endophytes GE 17-18
CK
Rb1/Rb2
β-Glucosidase
Sulfolobusacidocaldarius
CK
Rb1/Rb2/Rb3/Rc
β-Glucosidase
Aspergillusniger
CK
Rb1/Rb2
β-Glucosidase
Microbacteriuesteraromaticum
C-O
Rb2
β-Glucosidase
Cellulosimicrobiumcelluloses
C-Y
Rb2
β-Glucosidase
Terrabacter ginsenosidimutans
C-Mc
Rc
β-Glucosidase
Terrabacter ginsenosidimutans
C-Mc1
Rc
β-Glucosidase
Cellulosimicrobium cellulans
C-Mx
Rb3
β-Glucosidase
Terrabacter ginsenosidimutans
Rg2
Re
β-Glucosidase
Microbacterium esteraromaticum
Rg2
Re
β-Glucosidase
Mucilaginibacter
Rg2
Re
β-Glucosidase
Pseudonocardia
Rh1
Rg1
β-Glucosidase
Microbacterium esteraromaticum
Rh1
Rf
β-Glucosidase
Pyrococcus furiosus
Rh1
Rf
β-Glucosidase
Aspergillus niger
Rh1
Rg2
α-L-Rhamnosidase
Absidia
Rh1
R2
β-Xylosidase
Thermoanaerobacterium
F1
Rg1
β-Glucosidase
Fusarium moniliforme
F1
Rg1
β-Glucosidase
Penicillium sclerotiorum
F1
Rg1
β-Glucosidase
Sanguibacter keddieii
G17 : gypénoside XVII ; G75 : gypénoside LXXV ; C-O : composé O ; C-Y : composé Y ; C-Mc1 : composé Mc1 ; C-Mc : composé Mc ; C-Mx : composé Mx ; C-K : composé K.
Les groupes de sucres C-6 et C-20 des ginsénosides triols peuvent également être hydrolysés par les glycosides hydrolases. Le ginsénoside Rg2 peut être obtenu en hydrolysant le glucose C-20 dans la molécule Re par la glycosidase. La glucosidase recombinante clonée à partir de Microbacterium esteraromaticum, Mucillaginibacter et Pseudonocardia peut non seulement convertir le ginsénoside Re en Rg2, mais aussi le ginsénoside Rg1 en Rh1. Le glucose, le rhamnose et le xylose en dehors de la position C-6 des ginsénosides Rf, Rg2 et R2 peuvent tous être convertis pour préparer le Rh1. Contrairement au ginsénoside Rh1, le ginsénoside F1 n'a qu'un seul glucose attaché à la position C-20 de son aglycone. La glucosidase de Fusarium moniliforme, Penicillium sclerotiorum et Sanguibacter keddieii peut hydrolyser spécifiquement le glucose C-6 du ginsénoside Rg1 pour produire le ginsénoside F1.
La glycoside hydrolase n'est pas seulement utilisée pour transformer et préparer des ginsénosides rares et actifs, mais elle est également largement utilisée pour hydrolyser et modifier des saponines telles que la réglisse, le soja et l'igname (tableau 2). La glucuronidase isolée et purifiée à partir de Streptococcus LJ-22 et Penicillium purpurogenum Li-3 peut hydrolyser la glycyrrhizine pour produire de l'acide monoglucuronique glycyrrhizine, et il n'y a pas de sous-produit d'acide glycyrrhétinique. Morana et al. ont utilisé la glucuronidase dérivée d'Aspergillus niger pour hydrolyser complètement la glycyrrhizine et produire de l'acide glycyrrhétinique. La soyasaponine hydrolase isolée et purifiée d'Aspergillus oryzae peut hydrolyser la soyasaponine I pour produire le soyasaponol B. Une nouvelle saponine hydrolase de soja dans Neocosmospora vasinfecta peut convertir les saponines de soja I, II et III en saponine de soja B, ce qui constitue un outil efficace pour préparer la saponine de soja avec un effet anti-oxydant et une régulation des lipides sanguins. Parmi les saponines stéroïdiennes, la recherche et la comparaison de la modification par hydrolyse de la chaîne de sucre de la dioscine sont systématiques. Inoue et al. ont isolé et purifié une glucosidase de Costus speciosus qui peut hydrolyser la diosgénine originale pour produire de la diosgénine. Liu et al. ont isolé, purifié et cloné à partir d'Aspergillus oryzae pour obtenir une dioscine hydrolase recombinante, qui peut hydrolyser les groupes glucosyl et α-1,4 rhamnosyl de la dioscine pour produire de la dioscine III. L'α-L-rhamnosidase isolée et purifiée par Feng et al. à partir de Curvularia lunata peut hydrolyser le groupe α-1,2 rhamnosyl de la dioscine pour produire la dioscine V. Qian et al. ont isolé et purifié une α-L-rhamnosidase à partir de foie de bœuf frais, qui peut hydrolyser les groupes α-1,2 et α-1,4 rhamnosyl de la diosgénine pour former un groupe glucose. -Diosgénine. Fu et al. ont isolé et purifié la diosgénine hydrolase de l'Absidia, qui peut hydrolyser complètement la diosgénine en diosgénine.
Tableau 2. Biotransformation d'autres saponines par la glycosidase
Produit
Substrat
Réaction
Organisme
GAMG
Glycyrrhizine
β-Glucuronidase
Streptocoques
GAMG
Glycyrrhizine
β-Glucuronidase
Penicillium purpurogenum
Acide glycyrrhétinique
Glycyrrhizine
β-Glucuronidase
Aspergillus niger
Soyasapogénol B
Soyasaponine I
Saponine hydrolase de soja
Aspergillus oryzae
Soyasapogénol B
Soyasaponine
Saponine hydrolase de soja
Neocosmospora vasinfecta
Dioscin
Protodioscine
β-Glucosidase
Costus speciosus
Progénine III
Protodioscine
Protodioscine-glycosidase
Aspergillus oryzae
Progénine V
Dioscin
α-L-Rhamnosidase
Curvularia lunata
Diosgényl-glucoside
Dioscin
α-L-Rhamnosidase
Foie bovin
Diosgénine
Dioscin
Dioscin-glycosidase
Absidia
GAMG : Mono-glucuronide d'acide glycyrrhétique
Contactez-nous dès maintenant !
Si vous avez besoin de Price, veuillez indiquer vos coordonnées dans le formulaire ci-dessous. Nous vous contacterons généralement dans les 24 heures. Vous pouvez également m'envoyer un courriel info@longchangchemical.com pendant les heures de travail ( 8:30 am to 6:00 pm UTC+8 Mon.~Sat. ) ou utilisez le chat en direct du site web pour obtenir une réponse rapide.