Saponine sind eine Klasse von Glykosiden, bei denen die Aglykone Triterpen- oder Steranverbindungen sind. Sie sind einer der wirksamen Bestandteile vieler chinesischer pflanzlicher Arzneimittel wie Ginseng, Süßholz und Yamswurzel (die Hauptstruktur der Saponine ist in Abbildung 1 dargestellt). Stärkung der Immunität und anderer Funktionen. In der Literatur gibt es viele Berichte über die Biotransformation von Ginsenosiden. Gegenwärtig sind mehr als 150 Arten von Ginsenosiden getrennt und identifiziert worden. Der Gehalt der Ginsenoside Rb1, Rb2, Rc, Rd, Re und Rg1 beträgt bis zu 80%, während der Gehalt der Ginsenoside Rg3, Rh2, F2 und Compound K (C-K) und anderer seltener Saponine gering oder gar nicht vorhanden ist. Studien haben gezeigt, dass einige seltene Saponine gute pharmakologische Aktivitäten haben. Aufgrund ihres geringen Gehalts sind die Herstellung und Produktion jedoch eingeschränkt. Die gleiche Art von Ginsenosid hat das gleiche Aglykon, aber die Zuckerkette ist unterschiedlich. Seltene Ginsenoside und der höhere Gehalt der gleichen Saponinart unterscheiden sich oft nur durch 2 bis 3 Zuckergruppen. Daher kann dieselbe Art von aktivem seltenem Saponin durch enzymatische Hydrolyse von Saponin mit hohem Gehalt hergestellt werden.
Quick answer: A practical enzyme or food-ingredient decision starts with the process target, then checks activity, application window, sensory impact, and batch-to-batch consistency before scale-up.
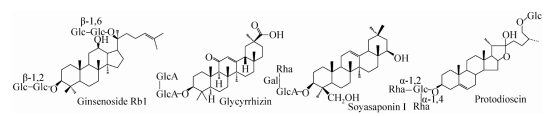
Abbildung 1. Struktur der wichtigsten Saponine
Verschiedene Glykosidhydrolasen haben unterschiedliche Selektivitäten, und auch die Wege zur Hydrolyse von Ginsenosiden sind unterschiedlich. Wie in Tabelle 1 dargestellt, können verschiedene Glykosidhydrolasen zur Herstellung verschiedener seltener Ginsenoside verwendet werden. Ginsenosid Rd kann durch Hydrolyse der äußeren C-20-Zuckergruppe der Ginsenoside Rb1, Rb2, Rb3 und Rc hergestellt werden. Die aus der chinesischen weißen Jadeschnecke und Thermus caldophilus isolierte und gereinigte β-Glucosidase kann das Ginsenosid Rb1 in Rd umwandeln. Kim et al. gewannen sie aus Bodenmikroorganismen mit Hilfe der molekularen Klonierungstechnologie, um Ginsenosid Rb1 umzuwandeln, das die rekombinante Glykosidhydrolase von Rd ist. Anschließend klonierten die Forscher Glucosidase aus Thermotoga thermarum und Bifidobacterium longum H-1, was die Effizienz der Transformation und der Herstellung von Ginsenosid Rd verbesserte. Die aus Flavobacterium johnsoniae und Thermus thermophilus durch rekombinante Technologie gewonnene Glucosidase kann nicht nur das Ginsenosid Rb1 in Rd umwandeln, sondern auch die C-20-Zuckerkette des Gypenosids XVII (G17) hydrolysieren, um das Ginsenosid F2 herzustellen. Zusätzlich zur Glucosidase wurde α-L-Arabinofuranosid-Hydrolase, die Ginsenosid Rc in Rd umwandeln kann, aus Ginsengwurzel und Leuconostoc sp. gewonnen. Α-L-Arabinofuranosid-Hydrolase und α-L-Arabinopyranosid-Hydrolase werden aus Bifidobacterium breve und Bifidobacterium longum gewonnen, die Ginsenosid Rc und Rb2 in Rd umwandeln können. In der Literatur wurde berichtet, dass die α-L-Arabinofuranosid-Hydrolase in Caldicellulosiruptor saccharolyticus und Rhodanobacter ginsenosidimutans nicht nur das Ginsenosid Rc zu Rd hydrolysieren, sondern auch die Verbindung Mc1 (C-Mc1) in F2 umwandeln kann. Die von Yu et al. aus Aspergillus isolierte und gereinigte Glykosidhydrolase kann alle Ginsenoside Rb1, Rb2, Rb3 und Rc in Rd umwandeln. Einige Glykosidhydrolasen können die Zuckerketten an der C-20-Position in Molekülen wie den glykolartigen Ginsenosiden Rb1, Rb2, Rb3, Rc und Rd vollständig hydrolysieren, um das Ginsenosid Rg3 zu erzeugen, das die Produktion von Rg3 in großem Maßstab ermöglicht und als Antitumormittel entwickelt wird. Die Glucosidase in Paecilomyces bainier und Microbacterium esteraromaticum kann das Ginsenosid Rb1 direkt zu Rg3 hydrolysieren, während die aus Microbacterium esteraromaticum isolierte und gereinigte Glucosidase das Ginsenosid Rb2 zu Rg3 hydrolysieren kann. Die rekombinante Glykosidhydrolase, die mit Hilfe der molekularen Klonierungstechnologie aus Pseudonocardia geklont wurde, kann die Ginsenoside Rb1, Rb3 und Rd zu Rg3 umwandeln. In ähnlicher Weise kann eine Reihe aktiver seltener Ginsenoside durch Hydrolyse der Zuckergruppe an Position C-3 in Ginsenosiden hergestellt werden. Die rekombinante Glucosidase, die aus Sphingomonas und Sphingopyxis alaskensis geklont wurde, kann die Glukose außerhalb der Zuckerkette an der Position C-3 in den Ginsenosidmolekülen Rb1, Rb2, Rc, Rd und Rg3 hydrolysieren und G17, Compound O (CO) und C-Mc1, F2 und Rh2 herstellen. Einige Glycosidasen können die innere Glucosylgruppe an der C-3-Position direkt hydrolysieren. Beispielsweise kann die Glucosidase aus Terrabacter ginsenosidimutans und Esteya vermicola die Zuckerkette an der C-3-Position der Ginsenosidmoleküle Rb1, Rb2, Rb3, Rc und Rd hydrolysieren, um das entsprechende Saponin LXXV (G75), Verbindung Y (C-Y), Verbindung Mx (C-Mx), Verbindung Mc (C-Mc) und C-K zu bilden. Darüber hinaus können einige Glykosidhydrolasen gleichzeitig die C-20- und C-3-Zuckergruppen in Ginsenosiden vom Glykol-Typ hydrolysieren. Die rekombinante Glucosidase, die aus Arthrobacter chlorophenolicus geklont wurde, kann die Ginsenoside Rb1, Rb2 und Rc in F2 umwandeln. Die Glykosidhydrolase in Fusobacterium K60, den endophytischen Pilzen GE 17-18, Sulfolobus acidocaldarius, Aspergillus niger und Microbacteriu esteraromaticum kann das Ginsenosid Rb1 hydrolysieren, um C-K herzustellen.
Tabelle 1. Biotransformation von Ginsenosiden durch Glycosidase
| Produkt | Substrat | Reaktion | Organismus |
| Rd | Rb1 | β-Glucosidase | China weiße Jadeschnecke |
| Rd | Rb1 | β-Glucosidase | Thermus Caldophilus |
| Rd | Rb1 | β-Glucosidase | Ungezüchtete Bakterien |
| Rd | Rb1 | β-Glucosidase | Thermotoga thermarum |
| Rd | Rb1 | β-Glucosidase | Bifidobacterium longum H-1 |
| Rd | Rb1 | β-Glucosidase | Flavobacterium johnsoniae |
| Rd | Rb1 | β-Glucosidase | Thermus thermophilus |
| Rd | Rb1 | β-Glucosidase | Penicillium oxalicum |
| Rd | Rb1 | β-Glucosidase | Cladosporium fulvum |
| Rd | Rc | α-L-Arabinofuranosidase | Panax Ginseng |
| Rd | Rc | α-L-Arabinofuranosidase | Leuconostoc |
| Rd | Rc | α-L-Arabinofuranosidase | Bifidobacterium breve |
| Rd | Rc | α-L-Arabinofuranosidase | Bifidobacterium longum |
| Rd | Rc | α-L-Arabinofuranosidase | Caldicellulosiruptor saccharolyticus |
| Rd | Rc | α-L-Arabinofuranosidase | Rhodanobacter Ginsenosidimutans |
| Rd | Rb2 | α-L-Arabinopyranosidase | Bifidobacterium breve |
| Rd | Rb2 | α-L-Arabinopyranosidase | Bifidobacterium longum |
| Rd | Rb1/Rb2/Rb3/Rc | Glykosidase | Aspergillus |
| Rg3 | Rb1 | β-Glucosidase | Paecilomyces bainier |
| Rg3 | Rb1 | β-Glucosidase | Mikrobakterium esteraromaticum |
| Rg3 | Rb2 | β-Glucosidase | Mikrobakterium esteraromaticum |
| Rg3 | Rb1/Rb3/Rd | β-Glucosidase | Pseudonokardie |
| G17 | Rb1 | β-Glucosidase | Sphingomonas |
| G17 | Rb1 | β-Glucosidase | Sphingopyxis alaskensis |
| G17 | Rb1 | β-Glucosidase | Cellulosimicrobium Zellulose |
| G75 | Rb1 | β-Glucosidase | Terrabacter Ginsenosidimutans |
| G75 | Rb1 | β-Glucosidase | Esteya vermicola |
| F2 | G17 | β-Glucosidase | Flavobacterium johnsoniae |
| F2 | G17 | β-Glucosidase | Thermus thermophilus |
| F2 | C-Mc1 | α-L-Arabinofuranosidase | Caldicellulosiruptor saccharolyticus |
| F2 | C-Mc1 | α-L-Arabinofuranosidase | Rhodanobacter Ginsenosidimutans |
| F2 | Rd | β-Glucosidase | Cellulosimicrobium Zellulose |
| F2 | Rb1/Rb2/Rc | β-Glucosidase | Arthrobacter chlorophenolicus |
| Rh2 | Rg3 | β-Glucosidase | Sphingopyxis alaskensis |
| CK | Rd | β-Glucosidase | Terrabacter Ginsenosidimutans |
| CK | Rd | β-Glucosidase | Esteya vermicola |
| CK | Rb1 | β-Glucosidase | Fusobacterium K-60 |
| CK | Rb1 | β-Glucosidase | endophytische Pilze GE 17-18 |
| CK | Rb1/Rb2 | β-Glucosidase | Sulfolobus acidocaldarius |
| CK | Rb1/Rb2/Rb3/Rc | β-Glucosidase | Aspergillus niger |
| CK | Rb1/Rb2 | β-Glucosidase | Microbacteriu esteraromaticum |
| C-O | Rb2 | β-Glucosidase | Cellulosimicrobium Zellulose |
| C-Y | Rb2 | β-Glucosidase | Terrabacter ginsenosidimutans |
| C-Mc | Rc | β-Glucosidase | Terrabacter ginsenosidimutans |
| C-Mc1 | Rc | β-Glucosidase | Cellulosimicrobium cellulans |
| C-Mx | Rb3 | β-Glucosidase | Terrabacter ginsenosidimutans |
| Rg2 | Re | β-Glucosidase | Mikrobakterium esteraromaticum |
| Rg2 | Re | β-Glucosidase | Mucilaginibacter |
| Rg2 | Re | β-Glucosidase | Pseudonokardie |
| Rh1 | Rg1 | β-Glucosidase | Mikrobakterium esteraromaticum |
| Rh1 | Rf | β-Glucosidase | Pyrococcus furiosus |
| Rh1 | Rf | β-Glucosidase | Aspergillus niger |
| Rh1 | Rg2 | α-L-Rhamnosidase | Absidia |
| Rh1 | R2 | β-Xylosidase | Thermoanaerobakterium |
| F1 | Rg1 | β-Glucosidase | Fusarium moniliforme |
| F1 | Rg1 | β-Glucosidase | Penicillium sclerotiorum |
| F1 | Rg1 | β-Glucosidase | Sanguibacter keddieii |
G17: Gypenosid XVII; G75: Gypenosid LXXV; C-O: Verbindung O; C-Y: Verbindung Y; C-Mc1: Verbindung Mc1; C-Mc: Verbindung Mc; C-Mx: Verbindung Mx; C-K: Verbindung K.
Die C-6- und C-20-Zuckergruppen in Triol-Ginsenosiden können auch durch Glykosidhydrolasen hydrolysiert werden. Ginsenosid Rg2 kann durch Hydrolyse von C-20-Glukose im Re-Molekül durch Glykosidase gewonnen werden. Die rekombinante Glucosidase, die aus Microbacterium esteraromaticum, Mucillaginibacter und Pseudonocardia geklont wurde, kann nicht nur Ginsenosid Re in Rg2 umwandeln, sondern auch Ginsenosid Rg1 in Rh1. Glucose, Rhamnose und Xylose außerhalb der C-6-Position von Ginsenosid Rf, Rg2 und R2 können alle zur Herstellung von Rh1 umgewandelt werden. Im Gegensatz zu Ginsenosid Rh1 ist bei Ginsenosid F1 nur eine Glukose an die C-20-Position seines Aglykons gebunden. Die Glucosidase in Fusarium moniliforme, Penicillium sclerotiorum und Sanguibacter keddieii kann die C-6-Glucose von Ginsenosid Rg1 spezifisch hydrolysieren, um Ginsenosid F1 herzustellen.
Glykosidhydrolase wird nicht nur zur Umwandlung und Herstellung aktiver seltener Ginsenoside verwendet, sondern auch zur Hydrolyse und Modifizierung von Saponinen wie Süßholz, Sojabohnen und Yamswurzel (Tabelle 2). Die aus Streptococcus LJ-22 und Penicillium purpurogenum Li-3 isolierte und gereinigte Glucuronidase kann Glycyrrhizin hydrolysieren, um Glycyrrhizin in Form von Monoglucuronsäure zu produzieren, und es entsteht kein Nebenprodukt Glycyrrhetinsäure. Morana et al. verwendeten Glucuronidase aus Aspergillus niger, um Glycyrrhizin vollständig zu hydrolysieren und Glycyrrhetinsäure zu erzeugen. Die aus Aspergillus oryzae isolierte und gereinigte Sojasaponin-Hydrolase kann Sojasaponin I hydrolysieren, um Sojasaponol B zu erzeugen. Eine neue Sojasaponin-Hydrolase in Neocosmospora vasinfecta kann Sojasaponin I, II und III in Sojasaponin B umwandeln, was ein wirksames Mittel zur Herstellung von Sojasaponin mit Antioxidationswirkung und Blutfettregulierung darstellt. Unter den Steroidsaponinen ist die Erforschung und der Vergleich der Hydrolysemodifikation der Zuckerkette von Dioscin systematisch. Inoue et al. isolierten und reinigten eine Glucosidase aus Costus speciosus, die das ursprüngliche Diosgenin hydrolysieren kann, um Diosgenin herzustellen. Liu et al. isolierten, reinigten und klonierten aus Aspergillus oryzae eine rekombinante Dioscin-Hydrolase, die die Glucosyl- und α-1,4-Rhamnosylgruppen in Dioscin hydrolysieren kann, um Dioscin III zu erzeugen. Die von Feng et al. aus Curvularia lunata isolierte und gereinigte α-L-Rhamnosidase kann die α-1,2-Rhamnosylgruppe in Dioscin unter Bildung von Dioscin V hydrolysieren. Qian et al. isolierten und reinigten eine α-L-Rhamnosidase aus frischer Rinderleber, die die beiden α-1,2- und α-1,4-Rhamnosylgruppen in Diosgenin unter Bildung einer Glucosegruppe hydrolysieren kann. -Diosgenin. Fu et al. isolierten und reinigten Diosgenin-Hydrolase aus Absidia, die Diosgenin vollständig zu Diosgenin hydrolysieren kann.
Tabelle 2. Biotransformation von anderen Saponinen durch Glycosidase
| Produkt | Substrat | Reaktion | Organismus |
| GAMG | Glycyrrhizin | β-Glucuronidase | Streptokokkus |
| GAMG | Glycyrrhizin | β-Glucuronidase | Penicillium purpurogenum |
| Glycyrrhetinsäure | Glycyrrhizin | β-Glucuronidase | Aspergillus niger |
| Sojasapogenol B | Sojasaponin I | Sojabohnen-Saponin-Hydrolase | Aspergillus oryzae |
| Sojasapogenol B | Sojasaponin | Sojabohnen-Saponin-Hydrolase | Neocosmospora vasinfecta |
| Dioscin | Protodioscin | β-Glucosidase | Costus speciosus |
| Progenin III | Protodioscin | Protodioscin-Glykosidase | Aspergillus oryzae |
| Progenin V | Dioscin | α-L-Rhamnosidase | Curvularia lunata |
| Diosgenyl-Glucosid | Dioscin | α-L-Rhamnosidase | Rinderleber |
| Diosgenin | Dioscin | Dioscin-Glykosidase | Absidia |
GAMG: Glycyrrhetinsäure-Mono-Glucuronid
A practical sourcing checklist for enzyme, biotech, and food-ingredient topics
In enzyme and food-processing projects, the most useful decision frame is usually application fit plus process stability: which ingredient performs under the intended pH, temperature, time, and substrate conditions without creating a downstream quality or compliance problem.
- Define the processing target first: flavor, hydrolysis, texture, fermentation, cleaning, and bioprocess applications often need very different activity profiles.
- Check the real operating window: pH, temperature, residence time, and substrate type often matter more than a headline product claim.
- Review consistency and downstream impact: dosage, sensory influence, filtration, and shelf-life behavior can all affect the final commercial value.
- Use pilot validation: small production tests usually reveal the most useful differences in activity, efficiency, and process fit.
Recommended product references
- Longzyme Lipase: A direct product reference for lipase-related food, cleaning, or bioprocess discussions.
- Longzyme Beta-Amylase: A practical enzyme reference when starch conversion and food-processing activity are under review.
- Longzyme Compound Glucoamylase: A useful enzyme reference when saccharification or related processing performance matters.
- Hefeextrakt: A practical ingredient reference when flavor, fermentation, or nutrient-support applications are involved.
FAQ for buyers and formulators
Why is a high-activity enzyme not automatically the best commercial choice?
Because the best enzyme is the one that performs reliably under the actual process conditions and gives the desired downstream result without creating new issues.
Should food and biotech ingredients be selected from data sheets alone?
It is usually safer to pair the specification review with a pilot or application test because real substrates and process windows can change the result a lot.
Kontaktieren Sie uns jetzt!
Wenn Sie einen Preis benötigen, tragen Sie bitte Ihre Kontaktdaten in das unten stehende Formular ein. Wir werden uns in der Regel innerhalb von 24 Stunden mit Ihnen in Verbindung setzen. Sie können mir auch mailen info@longchangchemical.com während der Geschäftszeiten ( 8:30 bis 18:00 Uhr UTC+8 Mo.~Sa. ) oder nutzen Sie den Live-Chat auf der Website, um eine schnelle Antwort zu erhalten.
| Verbindung Glucoamylase | 9032-08-0 |
| Pullulanase | 9075-68-7 |
| Xylanase | 37278-89-0 |
| Cellulase | 9012-54-8 |
| Naringinase | 9068-31-9 |
| β-Amylase | 9000-91-3 |
| Glukose-Oxidase | 9001-37-0 |
| alpha-Amylase | 9000-90-2 |
| Pektinase | 9032-75-1 |
| Peroxidase | 9003-99-0 |
| Lipase | 9001-62-1 |
| Katalase | 9001-05-2 |
| TANNASE | 9025-71-2 |
| Elastase | 39445-21-1 |
| Urease | 9002-13-5 |
| DEXTRANASE | 9025-70-1 |
| L-Milch-Dehydrogenase | 9001-60-9 |
| Dehydrogenase Malat | 9001-64-3 |
| Cholesterin-Oxidase | 9028-76-6 |