Saponin adalah kelas glikosida di mana aglikon adalah senyawa triterpen atau steran. Saponin adalah salah satu bahan yang efektif dari banyak obat herbal Cina seperti ginseng, licorice, dan ubi (struktur saponin utama ditunjukkan pada Gambar 1). Meningkatkan imunitas dan fungsi lainnya. Ada banyak laporan tentang biotransformasi ginsenosides dalam literatur. Saat ini, lebih dari 150 jenis ginsenosides telah dipisahkan dan diidentifikasi. Kandungan ginsenosides Rb1, Rb2, Rc, Rd, Re dan Rg1 setinggi 80%, sedangkan kandungan ginsenosides Rg3, Rh2, F2 dan Senyawa K (C-K) dan saponin langka lainnya memiliki kandungan yang sedikit atau tidak sama sekali. Penelitian telah menunjukkan bahwa beberapa saponin langka memiliki aktivitas farmakologis yang baik. Namun, karena kandungannya yang rendah, persiapan dan produksinya dibatasi. Jenis ginsenoside yang sama memiliki aglycon yang sama, tetapi rantai gulanya berbeda. Ginsenosides langka dan kandungan yang lebih tinggi dari jenis saponin yang sama seringkali hanya berbeda 2 hingga 3 gugus gula. Oleh karena itu, jenis saponin langka aktif yang sama dapat dibuat dengan hidrolisis enzimatik dari saponin berkadar tinggi.
Quick answer: A practical enzyme or food-ingredient decision starts with the process target, then checks activity, application window, sensory impact, and batch-to-batch consistency before scale-up.
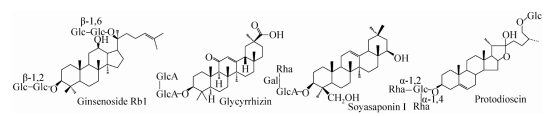
Gambar 1. Struktur saponin utama
Hidrolase glikosida yang berbeda memiliki selektivitas yang berbeda, dan jalur untuk menghidrolisis ginsenosida juga berbeda. Seperti yang ditunjukkan pada Tabel 1, hidrolase glikosida yang berbeda dapat digunakan untuk membuat ginsenosida langka yang berbeda. Ginsenoside Rd dapat dibuat dengan menghidrolisis ginsenosides Rb1, Rb2, Rb3 dan gugus gula luar C-20 Rc. β-glukosidase yang diisolasi dan dimurnikan dari siput giok putih Cina dan Thermus caldophilus dapat mengubah ginsenosida Rb1 menjadi Rd. Kim et al. memperolehnya dari mikroorganisme tanah dengan menggunakan teknologi kloning molekuler untuk mengkonversi ginsenosida Rb1 yang merupakan glikosida hidrolase rekombinan Rd. Selanjutnya, para peneliti mengkloning glukosidase dari Thermotoga thermarum dan Bifidobacterium longum H-1, yang meningkatkan efisiensi transformasi dan pembuatan ginsenosida Rd. Glukosidase yang diperoleh dari Flavobacterium johnsoniae dan Thermus thermophilus dengan teknologi rekombinan tidak hanya dapat mengubah ginsenoside Rb1 menjadi Rd, tetapi juga menghidrolisis rantai gula C-20 Gypenoside XVII (G17) untuk menghasilkan ginsenoside F2. Selain glukosidase, hidrolase α-L-arabinofuranosida yang dapat mengubah ginsenosida Rc menjadi Rd diperoleh dari akar ginseng dan Leuconostoc sp. Α-L-arabinofuranosida hidrolase dan hidrolase α-L-arabinopiranosida diperoleh dari Bifidobacterium breve dan Bifidobacterium longum, yang dapat mengubah ginsenosida Rc dan Rb2 menjadi Rd. Telah dilaporkan dalam literatur bahwa hidrolase α-L-arabinofuranosida dalam Caldicellulosiruptor saccharolyticus dan Rhodanobacter ginsenosidimutans tidak hanya dapat menghidrolisis ginsenosida Rc menjadi Rd, tetapi juga mengubah Senyawa Mc1 (C-Mc1) menjadi F2. Hidrolase glikosida yang diisolasi dan dimurnikan dari Aspergillus oleh Yu et al. dapat mengubah semua ginsenosida Rb1, Rb2, Rb3 dan Rc menjadi Rd. Beberapa hidrolase glikosida dapat sepenuhnya menghidrolisis rantai gula pada posisi C-20 dalam molekul seperti ginsenosida tipe glikol Rb1, Rb2, Rb3, Rc dan Rd, untuk menghasilkan ginsenosida Rg3, yang memungkinkan produksi Rg3 dalam skala besar dan dikembangkan sebagai obat agen anti tumor. Glukosidase dalam Paecilomyces bainier dan Microbacterium esteraromaticum dapat secara langsung menghidrolisis ginsenoside Rb1 menjadi Rg3, sedangkan glukosidase yang diisolasi dan dimurnikan dari Microbacterium esteraromaticum dapat menghidrolisis ginsenoside Rb2 menjadi Rg3. Glikosida hidrolase rekombinan yang dikloning dari Pseudonocardia dengan teknologi kloning molekuler dapat mengubah ginsenosida Rb1, Rb3 dan Rd untuk membuat Rg3. Demikian pula, serangkaian ginsenosida langka yang aktif dapat dibuat dengan menghidrolisis gugus gula pada posisi C-3 dalam ginsenosida. Glukosidase rekombinan yang dikloning dari Sphingomonas dan Sphingopyxis alaskensis dapat menghidrolisis glukosa di luar rantai gula pada posisi C-3 pada molekul ginsenosida Rb1, Rb2, Rc, Rd, dan Rg3, dan menyiapkan G17, Senyawa O (CO), serta C-Mc1, F2 dan Rh2. Beberapa glikosidase dapat secara langsung menghidrolisis gugus glukosil bagian dalam pada posisi C-3. Sebagai contoh, glukosidase dari Terrabacter ginsenosidimutans dan Esteya vermicola dapat menghidrolisis rantai gula pada posisi C-3 dari molekul ginsenosida Rb1, Rb2, Rb3, Rc dan Rd untuk menghasilkan saponin yang sesuai LXXV (G75), Senyawa Y (C-Y), Senyawa Mx (C-Mx), Senyawa Mc (C-Mc) dan C-K. Selain itu, beberapa hidrolase glikosida secara bersamaan dapat menghidrolisis gugus gula C-20 dan C-3 dalam ginsenosida tipe glikol. Glukosidase rekombinan yang dikloning dari Arthrobacter chlorophenolicus dapat mengubah ginsenosida Rb1, Rb2 dan Rc menjadi F2. Glikosida hidrolase pada Fusobacterium K60, jamur endofit GE 17-18, Sulfolobus acidocaldarius, Aspergillus niger dan Microbacteriu esteraromaticum dapat menghidrolisis ginsenosida Rb1 untuk menghasilkan C-K.
Tabel 1. Biotransformasi ginsenosida oleh glikosidase
| Produk | Substrat | Reaksi | Organisme |
| Rd | Rb1 | β-Glukosidase | Siput giok putih Cina |
| Rd | Rb1 | β-Glukosidase | Thermus caldophilus |
| Rd | Rb1 | β-Glukosidase | Bakteri yang tidak dikultur |
| Rd | Rb1 | β-Glukosidase | Thermotoga thermarum |
| Rd | Rb1 | β-Glukosidase | Bifidobacterium longum H-1 |
| Rd | Rb1 | β-Glukosidase | Flavobacterium johnsoniae |
| Rd | Rb1 | β-Glukosidase | Thermus thermophilus |
| Rd | Rb1 | β-Glukosidase | Penicillium oxalicum |
| Rd | Rb1 | β-Glukosidase | Cladosporium fulvum |
| Rd | Rc | Î ± -L-Arabinofuranosidase | Panax ginseng |
| Rd | Rc | Î ± -L-Arabinofuranosidase | Leuconostoc |
| Rd | Rc | Î ± -L-Arabinofuranosidase | Bifidobacterium singkat |
| Rd | Rc | Î ± -L-Arabinofuranosidase | Bifidobacterium longum |
| Rd | Rc | Î ± -L-Arabinofuranosidase | Caldicellulosiruptor saccharolyticus |
| Rd | Rc | Î ± -L-Arabinofuranosidase | Rhodanobacter ginsenosidimutans |
| Rd | Rb2 | α-L-Arabinopyranosidase | Bifidobacterium singkat |
| Rd | Rb2 | α-L-Arabinopyranosidase | Bifidobacterium longum |
| Rd | Rb1/Rb2/Rb3/Rc | Glikosidase | Aspergillus |
| Rg3 | Rb1 | β-Glukosidase | Paecilomyces bainier |
| Rg3 | Rb1 | β-Glukosidase | Mikrobakteri esteraromaticum |
| Rg3 | Rb2 | β-Glukosidase | Mikrobakteri esteraromaticum |
| Rg3 | Rb1/Rb3/Rd | β-Glukosidase | Pseudonokardia |
| G17 | Rb1 | β-Glukosidase | Sphingomonas |
| G17 | Rb1 | β-Glukosidase | Sphingopyxis alaskensis |
| G17 | Rb1 | β-Glukosidase | Cellulosimicrobium selulans |
| G75 | Rb1 | β-Glukosidase | Terrabacter ginsenosidimutans |
| G75 | Rb1 | β-Glukosidase | Esteya vermicola |
| F2 | G17 | β-Glukosidase | Flavobacterium johnsoniae |
| F2 | G17 | β-Glukosidase | Thermus thermophilus |
| F2 | C-Mc1 | Î ± -L-Arabinofuranosidase | Caldicellulosiruptor saccharolyticus |
| F2 | C-Mc1 | Î ± -L-Arabinofuranosidase | Rhodanobacter ginsenosidimutans |
| F2 | Rd | β-Glukosidase | Cellulosimicrobium selulans |
| F2 | Rb1 / Rb2 / Rc | β-Glukosidase | Arthrobacter chlorophenolicus |
| Rh2 | Rg3 | β-Glukosidase | Sphingopyxis alaskensis |
| CK | Rd | β-Glukosidase | Terrabacter ginsenosidimutans |
| CK | Rd | β-Glukosidase | Esteya vermicola |
| CK | Rb1 | β-Glukosidase | Fusobacterium K-60 |
| CK | Rb1 | β-Glukosidase | jamur endofit GE 17-18 |
| CK | Rb1/Rb2 | β-Glukosidase | Sulfolobus acidocaldarius |
| CK | Rb1/Rb2/Rb3/Rc | β-Glukosidase | Aspergillus niger |
| CK | Rb1/Rb2 | β-Glukosidase | Mikrobakteriu esteraromaticum |
| C-O | Rb2 | β-Glukosidase | Cellulosimicrobium selulans |
| C-Y | Rb2 | β-Glukosidase | Terrabacter ginsenosidimutans |
| C-Mc | Rc | β-Glukosidase | Terrabacter ginsenosidimutans |
| C-Mc1 | Rc | β-Glukosidase | Cellulosimicrobium cellulans |
| C-Mx | Rb3 | β-Glukosidase | Terrabacter ginsenosidimutans |
| Rg2 | Kembali | β-Glukosidase | Microbacterium esteraromaticum |
| Rg2 | Kembali | β-Glukosidase | Mucilaginibacter |
| Rg2 | Kembali | β-Glukosidase | Pseudonokardia |
| Rh1 | Rg1 | β-Glukosidase | Microbacterium esteraromaticum |
| Rh1 | Rf | β-Glukosidase | Pyrococcus furiosus |
| Rh1 | Rf | β-Glukosidase | Aspergillus niger |
| Rh1 | Rg2 | Î ± -L-Rhamnosidase | Absidia |
| Rh1 | R2 | β-Xilosidase | Thermoanaerobacterium |
| F1 | Rg1 | β-Glukosidase | Fusarium moniliforme |
| F1 | Rg1 | β-Glukosidase | Penicillium sclerotiorum |
| F1 | Rg1 | β-Glukosidase | Sanguibacter keddieii |
G17: gypenoside XVII; G75: gypenoside LXXV; C-O: senyawa O; C-Y: senyawa Y; C-Mc1: senyawa Mc1; C-Mc: senyawa Mc; C-Mx: senyawa Mx; C-K: senyawa K.
Gugus gula C-6 dan C-20 dalam triol ginsenosida juga dapat dihidrolisis oleh hidrolase glikosida. Ginsenoside Rg2 dapat diperoleh dengan menghidrolisis glukosa C-20 dalam molekul Re oleh glikosidase. Glukosidase rekombinan yang dikloning dari Microbacterium esteraromaticum, Mucillaginibacter dan Pseudonocardia tidak hanya dapat mengubah ginsenoside Re menjadi Rg2, tetapi juga Ginsenoside Rg1 diubah menjadi Rh1. Glukosa, rhamnosa dan xilosa di luar posisi C-6 dari ginsenoside Rf, Rg2 dan R2 semuanya dapat dikonversi untuk menyiapkan Rh1. Tidak seperti ginsenosida Rh1, ginsenosida F1 hanya memiliki satu glukosa yang melekat pada posisi C-20 dari aglikonnya. Glukosidase pada Fusarium moniliforme, Penicillium sclerotiorum dan Sanguibacter keddieii secara khusus dapat menghidrolisis glukosa C-6 dari ginsenoside Rg1 untuk menghasilkan ginsenoside F1.
Glikosida hidrolase tidak hanya digunakan untuk mengubah dan membuat ginsenosida langka yang aktif, tetapi juga digunakan secara luas untuk menghidrolisis dan memodifikasi saponin seperti licorice, kedelai, dan ubi (Tabel 2). Glukuronidase yang diisolasi dan dimurnikan dari Streptococcus LJ-22 dan Penicillium purpurogenum Li-3 dapat menghidrolisis glikyrrhizin untuk menghasilkan asam monoglukuronat glikyrrhizin, dan tidak ada produk sampingan asam glikyrrhetinat. Morana dkk. menggunakan glukuronidase yang berasal dari Aspergillus niger untuk menghidrolisis glikyrrhizin sepenuhnya untuk menghasilkan asam glikyrrhetinat. Hidrolase saponin kedelai yang diisolasi dan dimurnikan dari Aspergillus oryzae dapat menghidrolisis soyasaponin I untuk menghasilkan soyasaponol B. Hidrolase saponin kedelai baru di Neocosmospora vasinfecta dapat mengubah saponin kedelai I, II, dan III menjadi saponin kedelai B, yang menyediakan alat yang efektif untuk menyiapkan saponin kedelai dengan anti oksidasi dan regulasi lipid darah. Di antara saponin steroid, penelitian dan perbandingan modifikasi hidrolisis rantai gula dioscin bersifat sistematis. Inoue dkk. mengisolasi dan memurnikan glukosidase dari Costus speciosus yang dapat menghidrolisis diosgenin asli untuk menghasilkan diosgenin. Liu dkk. mengisolasi, memurnikan dan mengkloning dari Aspergillus oryzae untuk mendapatkan dioscin hidrolase rekombinan, yang dapat menghidrolisis gugus glukosil dan α-1,4 rhamnosil pada dioscin untuk menghasilkan dioscin III. α-L-rhamnosidase yang diisolasi dan dimurnikan oleh Feng et al. dari Curvularia lunata dapat menghidrolisis gugus α-1,2 rhamnosil dalam dioscin untuk menghasilkan dioscin V. Qian dkk. mengisolasi dan memurnikan α-L-rhamnosidase dari hati sapi segar, yang dapat menghidrolisis dua gugus α-1,2 dan α-1,4 rhamnosil dalam diosgenin untuk membentuk gugus glukosa. -Diosgenin. Fu dkk. mengisolasi dan memurnikan diosgenin hidrolase dari Absidia, yang dapat menghidrolisis diosgenin sepenuhnya menjadi diosgenin.
Tabel 2. Biotransformasi saponin lain oleh glikosidase
| Produk | Substrat | Reaksi | Organisme |
| GAMG | Glycyrrhizin | β-Glukuronidase | Streptococcus |
| GAMG | Glycyrrhizin | β-Glukuronidase | Penicillium purpurogenum |
| Asam glikyrrhetinat | Glycyrrhizin | β-Glukuronidase | Aspergillus niger |
| Soyasapogenol B | Soyasaponin I | Hidrolase saponin kedelai | Aspergillus oryzae |
| Soyasapogenol B | Soyasaponin | Hidrolase saponin kedelai | Neocosmospora vasinfecta |
| Dioscin | Protodioscin | β-Glukosidase | Costus speciosus |
| Progenin III | Protodioscin | Protodioscin-glikosidase | Aspergillus oryzae |
| Progenin V | Dioscin | Î ± -L-Rhamnosidase | Curvularia lunata |
| Diosgenyl-glukosida | Dioscin | Î ± -L-Rhamnosidase | Hati sapi |
| Diosgenin | Dioscin | Dioscin-glikosidase | Absidia |
GAMG: Asam glukuronida mono-glukuronida
A practical sourcing checklist for enzyme, biotech, and food-ingredient topics
In enzyme and food-processing projects, the most useful decision frame is usually application fit plus process stability: which ingredient performs under the intended pH, temperature, time, and substrate conditions without creating a downstream quality or compliance problem.
- Define the processing target first: flavor, hydrolysis, texture, fermentation, cleaning, and bioprocess applications often need very different activity profiles.
- Check the real operating window: pH, temperature, residence time, and substrate type often matter more than a headline product claim.
- Review consistency and downstream impact: dosage, sensory influence, filtration, and shelf-life behavior can all affect the final commercial value.
- Use pilot validation: small production tests usually reveal the most useful differences in activity, efficiency, and process fit.
Recommended product references
- Longzyme Lipase: A direct product reference for lipase-related food, cleaning, or bioprocess discussions.
- Longzyme Beta-Amylase: A practical enzyme reference when starch conversion and food-processing activity are under review.
- Longzyme Compound Glucoamylase: A useful enzyme reference when saccharification or related processing performance matters.
- Ekstrak Ragi: A practical ingredient reference when flavor, fermentation, or nutrient-support applications are involved.
FAQ for buyers and formulators
Why is a high-activity enzyme not automatically the best commercial choice?
Because the best enzyme is the one that performs reliably under the actual process conditions and gives the desired downstream result without creating new issues.
Should food and biotech ingredients be selected from data sheets alone?
It is usually safer to pair the specification review with a pilot or application test because real substrates and process windows can change the result a lot.
Hubungi Kami Sekarang!
Jika Anda membutuhkan Harga, silakan isi informasi kontak Anda di formulir di bawah ini, kami biasanya akan menghubungi Anda dalam waktu 24 jam. Anda juga bisa mengirim email kepada saya info@longchangchemical.com selama jam kerja (8:30 pagi hingga 6:00 sore UTC+8 Senin-Sabtu) atau gunakan obrolan langsung situs web untuk mendapatkan balasan secepatnya.
| Senyawa Glukoamilase | 9032-08-0 |
| Pullulanase | 9075-68-7 |
| Xilanase | 37278-89-0 |
| Selulase | 9012-54-8 |
| Naringinase | 9068-31-9 |
| β-Amilase | 9000-91-3 |
| Glukosa oksidase | 9001-37-0 |
| alfa-Amilase | 9000-90-2 |
| Pektinase | 9032-75-1 |
| Peroksidase | 9003-99-0 |
| Lipase | 9001-62-1 |
| Katalase | 9001-05-2 |
| TANNASE | 9025-71-2 |
| Elastase | 39445-21-1 |
| Urease | 9002-13-5 |
| DEXTRANASE | 9025-70-1 |
| L-Laktat dehidrogenase | 9001-60-9 |
| Dehidrogenase malat | 9001-64-3 |
| Kolesterol oksidase | 9028-76-6 |