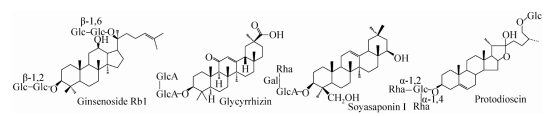
Saponinen zijn een klasse glycosiden waarvan de aglyconen triterpeen- of steraanverbindingen zijn. Ze zijn een van de effectieve ingrediënten van veel Chinese kruidengeneesmiddelen zoals ginseng, zoethout en yam (de hoofdstructuur van saponinen is weergegeven in figuur 1). Versterken immuniteit en andere functies. In de literatuur zijn veel rapporten te vinden over de biotransformatie van ginsenosiden. Op dit moment zijn er meer dan 150 soorten ginsenosiden gescheiden en geïdentificeerd. De inhoud van de ginsenosiden Rb1, Rb2, Rc, Rd, Re en Rg1 is zo hoog als 80%, terwijl de inhoud van de ginsenosiden Rg3, Rh2, F2 en Compound K (C-K) en andere zeldzame saponinen weinig of geen inhoud hebben. Studies hebben aangetoond dat sommige zeldzame saponinen goede farmacologische activiteiten hebben. Vanwege het lage gehalte is de bereiding en productie echter beperkt. Hetzelfde type ginsenoside heeft hetzelfde aglycon, maar de suikerketen is verschillend. Zeldzame ginsenosiden en het hogere gehalte aan dezelfde soort saponinen verschillen vaak slechts 2 tot 3 suikergroepen. Daarom kan hetzelfde type actieve zeldzame saponine worden bereid door enzymatische hydrolyse van saponine met een hoog gehalte.
Figuur 1. Structuur van de belangrijkste saponinen
Verschillende glycoside hydrolases hebben verschillende selectiviteiten en de routes om ginsenosiden te hydrolyseren zijn ook verschillend. Zoals tabel 1 laat zien, kunnen verschillende glycosidehydrolases worden gebruikt om verschillende zeldzame ginsenosiden te bereiden. Ginsenoside Rd kan worden bereid door hydrolyse van ginsenosiden Rb1, Rb2, Rb3 en de C-20 buitenste suikergroep van Rc. Het β-glucosidase geïsoleerd en gezuiverd uit de China white jade slak en Thermus caldophilus kan ginsenoside Rb1 omzetten in Rd. Kim et al. verkregen het van bodemmicro-organismen met behulp van moleculaire kloneringstechnologie om ginsenoside Rb1 om te zetten, wat het recombinante glycosidehydrolase van Rd is. Vervolgens kloonden onderzoekers glucosidase uit Thermotoga thermarum en Bifidobacterium longum H-1, wat de efficiëntie van de transformatie en bereiding van ginsenoside Rd verbeterde. De glucosidase verkregen uit Flavobacterium johnsoniae en Thermus thermophilus door middel van recombinante technologie kan niet alleen ginsenoside Rb1 omzetten in Rd, maar ook de C-20 suikerketen van Gypenoside XVII (G17) hydrolyseren om ginsenoside F2 te produceren. Naast glucosidase werd α-L-arabinofuranoside hydrolase, dat ginsenoside Rc kan omzetten in Rd, verkregen uit ginsengwortel en Leuconostoc sp. Α-L-arabinofuranoside hydrolase en α-L-arabinopyranoside hydrolase worden verkregen uit Bifidobacterium breve en Bifidobacterium longum, die ginsenoside Rc en Rb2 kunnen omzetten in Rd. In de literatuur is gemeld dat het α-L-arabinofuranoside hydrolase in Caldicellulosiruptor saccharolyticus en Rhodanobacter ginsenosidimutans niet alleen ginsenoside Rc kan hydrolyseren tot Rd, maar ook Compound Mc1 (C-Mc1) kan omzetten in F2. Het door Yu et al. uit Aspergillus geïsoleerde en gezuiverde glycosidehydrolase kan alle ginsenosiden Rb1, Rb2, Rb3 en Rc omzetten in Rd. Sommige glycosidehydrolases kunnen de suikerketens op de C-20 positie in moleculen zoals glycol-type ginsenosiden Rb1, Rb2, Rb3, Rc en Rd volledig hydrolyseren om ginsenoside Rg3 te genereren, wat grootschalige productie van Rg3 mogelijk maakt en ontwikkeld is als een geneesmiddel tegen tumoren. De glucosidase in Paecilomyces bainier en Microbacterium esteraromaticum kan ginsenoside Rb1 direct hydrolyseren in Rg3, terwijl de glucosidase geïsoleerd en gezuiverd uit Microbacterium esteraromaticum ginsenoside Rb2 kan hydrolyseren in Rg3. Het recombinante glycoside hydrolase gekloond uit Pseudonocardia door middel van moleculaire kloneringstechnologie kan de ginsenosiden Rb1, Rb3 en Rd omzetten in Rg3. Op dezelfde manier kan een serie actieve zeldzame ginsenosiden worden bereid door de suikergroep op positie C-3 in ginsenosiden te hydrolyseren. De recombinante glucosidase gekloond uit Sphingomonas en Sphingopyxis alaskensis kan de glucose hydrolyseren buiten de suikerketen op positie C-3 in de ginsenoside Rb1, Rb2, Rc, Rd, en Rg3 moleculen, en G17, Compound O (CO), en C-Mc1, F2 en Rh2 bereiden. Sommige glycosidasen kunnen de binnenste glucosylgroep op de C-3 positie direct hydrolyseren. Bijvoorbeeld, de glucosidase van Terrabacter ginsenosidimutans en Esteya vermicola kan de suikerketen op de C-3 positie van de ginsenoside Rb1, Rb2, Rb3, Rc en Rd moleculen hydrolyseren om de corresponderende saponine LXXV (G75), Compound Y (C-Y), Compound Mx (C-Mx), Compound Mc (C-Mc) en C-K te produceren. Daarnaast kunnen sommige glycosidehydrolases tegelijkertijd de C-20 en C-3 suikergroepen in ginsenosiden van het glycol-type hydrolyseren. Het recombinante glucosidase gekloond uit Arthrobacter chlorophenolicus kan ginsenosiden Rb1, Rb2 en Rc omzetten in F2. De glycoside hydrolase in Fusobacterium K60, endofytische schimmels GE 17-18, Sulfolobus acidocaldarius, Aspergillus niger en Microbacteriu esteraromaticum kan ginsenoside Rb1 hydrolyseren om C-K te produceren.
Tabel 1. Biotransformatie van ginsenosiden door glycosidase
De C-6 en C-20 suikergroepen in triol ginsenosiden kunnen ook gehydrolyseerd worden door glycoside hydrolases. Ginsenoside Rg2 kan worden verkregen door hydrolyse van C-20 glucose in het Re-molecuul door glycosidase. Het recombinante glucosidase gekloond uit Microbacterium esteraromaticum, Mucillaginibacter en Pseudonocardia kan niet alleen ginsenoside Re omzetten in Rg2, maar ook ginsenoside Rg1 wordt omgezet in Rh1. Glucose, rhamnose en xylose buiten de C-6 positie van ginsenoside Rf, Rg2 en R2 kunnen allemaal worden omgezet in Rh1. In tegenstelling tot ginsenoside Rh1 heeft ginsenoside F1 slechts één glucose gebonden aan de C-20 positie van zijn aglycon. Glucosidase in Fusarium moniliforme, Penicillium sclerotiorum en Sanguibacter keddieii kan specifiek de C-6 glucose van ginsenoside Rg1 hydrolyseren om ginsenoside F1 te produceren.
Glycosidehydrolase wordt niet alleen gebruikt om actieve zeldzame ginsenosiden te transformeren en te bereiden, maar wordt ook veel gebruikt om saponinen zoals zoethout, soja en yam te hydrolyseren en te modificeren (tabel 2). Glucuronidase geïsoleerd en gezuiverd uit Streptococcus LJ-22 en Penicillium purpurogenum Li-3 kan glycyrrhizine hydrolyseren om monoglucuronzuur glycyrrhizine te produceren, en er is geen bijproduct glycyrrhetinezuur. Morana et al. gebruikten glucuronidase afkomstig van Aspergillus niger om glycyrrhizine volledig te hydrolyseren en glycyrrhetinezuur te produceren. Sojasaponinehydrolase geïsoleerd en gezuiverd uit Aspergillus oryzae kan sojasaponine I hydrolyseren om sojasaponol B te produceren. Een nieuw sojasaponinehydrolase in Neocosmospora vasinfecta kan sojasaponine I, II en III omzetten in sojasaponine B, wat een effectief hulpmiddel is voor het bereiden van sojasaponine met antioxidatie en bloedlipidenregulatie. Van de steroïde saponinen is het onderzoek en de vergelijking van de hydrolysemodificatie van de suikerketen van dioscine systematisch. Inoue et al. isoleerden en zuiverden een glucosidase uit Costus speciosus die de oorspronkelijke diosgenine kan hydrolyseren om diosgenine te produceren. Liu et al. isoleerden, zuiverden en kloneerden uit Aspergillus oryzae een recombinant dioscine hydrolase, dat de glucosyl- en α-1,4-rhamnosylgroepen in dioscine kan hydrolyseren om dioscine III te produceren. De α-L-rhamnosidase geïsoleerd en gezuiverd door Feng et al. uit Curvularia lunata kan de α-1,2 rhamnosylgroep in dioscine hydrolyseren om dioscine V te produceren. Qian et al. isoleerden en zuiverden een α-L-rhamnosidase uit verse runderlever, die de twee α-1,2 en α-1,4 rhamnosylgroepen in diosgenine kan hydrolyseren om een glucosegroep te vormen. -Diosgenine. Fu et al. isoleerden en zuiverden diosgeninehydrolase uit Absidia, dat diosgenine volledig kan hydrolyseren tot diosgenine.
Tabel 2. Biotransformatie van andere saponinen door glycosidase
Product
Substraat
Reactie
Organisme
GAMG
Glycyrrhizine
β-glucuronidase
Streptococcus
GAMG
Glycyrrhizine
β-glucuronidase
Penicillium purpurogenum
Glycyrrhetinezuur
Glycyrrhizine
β-glucuronidase
Aspergillus niger
Sojasapogenol B
Sojasaponine I
Sojaboon saponine hydrolase
Aspergillus oryzae
Sojasapogenol B
Sojasaponine
Sojaboon saponine hydrolase
Neocosmospora vasinfecta
Dioscin
Protodioscine
β-Glucosidase
Costus speciosus
Progenine III
Protodioscine
Protodioscine-glycosidase
Aspergillus oryzae
Progenine V
Dioscin
α-L-Rhamnosidase
Curvularia lunata
Diosgenyl-glucoside
Dioscin
α-L-Rhamnosidase
Runderlever
Diosgenine
Dioscin
Dioscine-glycosidase
Absidia
GAMG: Glycyrrhetic zuur mono-glucuronide
Neem nu contact met ons op!
Als je Price nodig hebt, vul dan je contactgegevens in op het formulier hieronder. We nemen dan meestal binnen 24 uur contact met je op. Je kunt me ook een e-mail sturen info@longchangchemical.com tijdens kantooruren (8:30 tot 18:00 UTC+8 ma. ~ za.) of gebruik de live chat op de website voor een snel antwoord.